Агматин
| Агматин | |
|---|---|
 | |
| Общие | |
| Систематическое наименование |
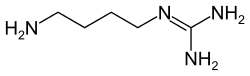
1-(4-аминобутил)гуанидин |
| Хим. формула | (NH-C(NH2)NH(CH2)4(NH2)) |
| Рац. формула | C5H14N4 |
| Физические свойства | |
| Молярная масса | 130.196 г/моль |
| Плотность | 1.2 г/мл |
| Термические свойства | |
| Температура | |
| • плавления | 102 °C |
| • кипения | 281 °C |
| Химические свойства | |
| Растворимость | |
| • в воде | высокая |
| Классификация | |
| Рег. номер CAS | 306-60-5 |
| PubChem | 199 |
| Рег. номер EINECS | 206-187-7 |
| SMILES | |
| InChI | |
| ChEBI | 17431 |
| ChemSpider | 194 |
| Приведены данные для стандартных условий (25 °C, 100 кПа), если не указано иное. | |
Агматин - это химическое вещество, образующееся в естественных условиях при декарбоксилировании аминокислоты аргинина. Было показано, что агматин оказывает модулирующее действие на множество молекулярных мишеней, в частности: нейромедиаторные системы, ионные каналы, синтез оксида азота (NO) и метаболизм полиаминов, это дает возможности для дальнейших исследований применения в фармакологии.
История
Соединение открыто в 1910 году Альбрехтом Косселем. На исследование биологической активности вещества ушло более 100 лет. Через год после его открытия было обнаружено, что Агматин может увеличивать кровоток у кроликов, однако физиологическая значимость этих результатов была поставлена под сомнение, учитывая требуемые высокие концентрации. В 1920-х годах исследователи из диабетической клиники Оскара Минковского показали, что агматин может оказывать умеренное гипогликемическое действие. В 1994 году произошло открытие эндогенного синтеза агматина у млекопитающих.
Пути метаболизма
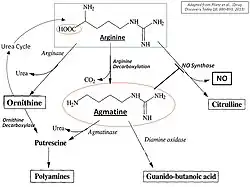
Биосинтез агматина путем декарбоксилирования аргинина конкурирует с другими аргининзависимыми путями, а именно: метаболизмом азота (цикл мочевины) и синтезом полиамина и оксида азота (NO). Разложение агматина происходит главным образом путем гидролиза, катализируемого агматиназой в мочевину и путресцин, диаминовый предшественник биосинтеза полиамина. Альтернативный путь(в основном, в периферических тканях) - катализируемое диаминоксидазой окисление в агматин-альдегид, который далее преобразуется альдегиддегидрогеназой в гуанидинобутират и секретируется почками.
Биологическая активность
Агматин блокирует катионные каналы, серотониновый 5-HT3 и н-холинорецептор, АТФ-чувствительные калиевые каналы, вольтаж-зависимые кальциевых каналы. Также, является агонистом имидазолиновых I1 и I2 рецепторов, связывается с α2-адренергическими, и серотониновыми 5-HT2A рецепторами.
Агматин обсуждался как предполагаемый нейромедиатор. Он синтезируется в головном мозге, хранится в синаптических пузырьках, высвобождается при деполяризации мембран и инактивируется агматиназой. Если не считать определения специфических ("собственных") постсинаптических рецепторов, агматин фактически соответствует критериям Генри Дейла для нейромедиатора, и может считаться нейромодулятором и ко-трансмиттером. Теоретическое существование агматинергических опосредованных нейрональных систем еще не доказано, хотя существование таких рецепторов подразумевается их значимостью в опосредовании как центральной, так и периферической нервной систем. Продолжаются поиски специфичных к агматину рецепторов и путей передачи.
Благодаря своей способности проходить через открытые катионные каналы агматин использовался в качестве суррогатной метрики возбудимости ионного канала в нервной ткани. При внешней стимуляции инкубируемой в агматине нервной ткани только клетки с открытыми каналами будут заполняться агматином, что позволяет идентифицировать, какие клетки чувствительны к этим стимулам и в какой степени они открывают свои катионные каналы в период стимуляции.
Системный приём агматина может потенцировать опиоидную анальгезию и предотвращать толерантность при длительном введении морфина у лабораторных грызунов. Накопленные данные убедительно свидетельствуют о том, что агматин подавляет опиоидную зависимость и рецидивы у нескольких видов животных.
Агматин производит умеренное снижение частоты сердечных сокращений и кровяного давления, по-видимому, активируя как центральную, так и периферическую системы управления через модуляцию нескольких своих молекулярных мишеней, включая подтипы имидазолиновых рецепторов, высвобождение норадреналина и выработку NO.
Инъекция сульфата агматина может увеличить потребление богатой углеводами пищи у сытых, но не у голодных крыс, этот эффект может быть опосредован нейропептидом Y. Однако, добавление вещества в питьевую воду приводит к снижению потребления воды и прироста массы тела. Гипогликемические эффекты агматина являются результатом одновременной модуляции нескольких молекулярных механизмов, участвующих в регуляции уровня глюкозы в крови.
Было показано, что агматин повышает скорость клубочковой фильтрации (СКФ) и оказывает нефропротекторное действие.