Allomerus decemarticulatus
| Allomerus decemarticulatus | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 Голова рабочего муравья | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Hymenopterida Отряд: Подотряд: Инфраотряд: Надсемейство: Семейство: Подсемейство: Триба: Род: Вид: Allomerus decemarticulatus |
||||||||||
| Международное научное название | ||||||||||
| Allomerus decemarticulatus Mayr, 1878[1] | ||||||||||
| ||||||||||
Allomerus decemarticulatus (лат.) — вид муравьёв из подсемейства мирмицины (Myrmicinae). Эндемики дождевых лесов Амазонии тропической Южной Америки. Обитает на растениях вида Hirtella physophora. Мелкие желтовато-коричневые муравьи (рабочие около 2 мм, матки около 5 мм). Этот вид отличается сложным и экстремальным хищническим поведением рабочих, которое включает симбиоз с растением и грибком. Они живут в карманах листьев растения-хозяина, Hirtella physophora. Эти листовые карманы представляют собой участки внутри растения между листьями и стеблем. Каждая колония, состоящая примерно из 1200 рабочих, обитает на одном дереве; однако муравьи рассредоточены по листовым карманам, и обычно на один карман приходится 40 рабочих.
Муравьи используют разнообразную пищу. Их рацион состоит в основном из крупных насекомых, которых они ловят на растении, но они также питаются некоторыми видами пищевых тел, производимых растением, и его нектаром. Для захвата добычи, которая гораздо крупнее их самих, они сооружают платформу с отверстиями на стебле растения (отверстия немного шире собственных тел муравьёв), которая служит ловушкой для ничего не подозревающей добычи. Для укрепления ловушки муравьи используют срезаемые волоски (трихомы) и собранную плесень, которая прорастает между трихомами и вокруг отверстий, заполняя и скрепляя структуру. Муравьи прячутся в ловушке и нападают, когда на неё садится любое прилетающее насекомое. Эта техника является примером хищничества из засады[2].
Описание

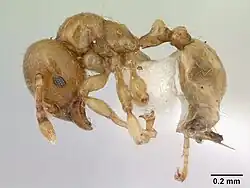
Мелкие желтовато-коричневые муравьи (рабочие около 2 мм). Антенны рабочих 10-сегментные, членики не достигают заднебокового угла головы, дистальная половина утолщена. Заднегрудной шов неглубокий и широкий. Проподеум не вооружён, дорсальная поверхность почти отсутствует, за ней следует наклонная поверхность, слабо угловатая между поверхностями. Проподеальное дыхальце маленькое. Петиоль со стебельком и с узлом хорошо выражен, узел короче стебелька. Петиолярный передневентральный шип хорошо развит. Бока мезоплевр и проподеума с несколькими продольными короткими и неправильными килями. Волоски обильные на дорсальной стороне мезосомы. Длинные волоски (более 0,13 мм длиной): шесть на передней стороне головы, около вершинного края и отстоящие; два ряда из нескольких волосков на лбу; несколько на промезонотуме, ни одного на проподеуме, несколько на петиоле, постпетиоле и брюшке. Короткие волоски (менее 0,07 мм) на всей дорсальной стороне мезосомы, на проподеуме их немного. Ряд коротких волосков на переднем крае наличника, центральный самый длинный, все выступают над наличником. Антенны и ноги с прижатым опушением. Основная окраска тела коричневато-жёлтая, волоски беловатые[1].
Общая длина тела королевы — 5,53 мм. Голова длиннее ширины, глаза выступающие. Антенны маток 11-сегментные, без выраженной булавы. Скутеллюм образует небольшой выступ с выпуклой дорсальной частью при виде сбоку. Проподеум угловатый между гранями. Проподеальное дыхальце открыто латерально. Петиолярный узел короче стебелька; они широко срастаются. Передневентральный отросток петиоля острый, без зубца или шипа. Тело гладкое и полированное. Голова с продольными полосами, особенно на задней части. Продольные бороздки на задней части мезоскутума. Аксилярная поперечная полоса и скутеллюм с густой и заметной продольной полосой. Проподеум с ограниченной полосой, косо продольной под проподеальным отростком. Волоски на теле обильные (длина волосков меньше ширины усиков), кроме большей части мезоскутума, анепистернума и катепистернума. Окраска одноцветная коричневая с чёрными глазами и аксилами[1].
Allomerus decemarticulatus близок к виду Allomerus brevipilosus, так как у их рабочих также 10-члениковые усики, пронотальный шов отсутствует, а проподеум округлый или слабо угловатый. Но от которого A. decemarticulatus отличается следующими признаками: наличник гладкий и блестящий; волоски на теле более длинные, самые длинные волоски около 0,12 мм; передние волоски на наличнике более длинные, центральный волосок заметный (у A. brevipilosus наличник с несколькими короткими продольными бороздками; волоски на теле короткие, самые длинные волоски около 0,07 мм; передние волоски на наличнике очень короткие, центральный волосок малозаметен)[1].
Мутуализм с растениями
Отношения между муравьями A. decemarticulatus и растениями Hirtella physophora очень специализированные. H. physophora — небольшое дерево в тропических лесах Амазонки, обитающее в подлеске. На одну особь H. physophora приходилось в среднем 1185±328 рабочих муравьёв, что составляло 39,2±5,2 рабочих на лист (от 19 до 36 листьев на растении), колонии всегда были моногинными (включали одну муравьиную матку)[2]. Это также пример мирмекофита, поскольку в нём есть муравьиные домики-домации, листовые мешочки и полые камеры, в которых постоянно гнездятся муравьи. Листовые мешочки образуются, когда край листа, прилегающий к черешку, загибается под него. В результате по обе стороны от стебля листа образуются две полые сферы. Муравьи живут внутри этих домаций. На одном дереве может быть только одна колония, но эта колония делится на множество разных домаций. В мешочках листьев также расположены внецветковые (экстрафлоральные) нектарники, которые обеспечивают муравьёв нектаром, производимым растением, а также некоторые пищевые тела, которые обеспечивают муравьёв питанием по мере их взросления и превращения в фуражиров и охотников на насекомых. Это первый известный случай, когда внецветковые нектарники наблюдаются внутри муравьиных домаций[3].
В некоторой степени это выглядит как паразитизм муравьёв, как будто они просто используют растение в качестве пищи и укрытия. Однако растение также получает большую выгоду от этих отношений. С помощью своих ловушек и хищничества муравьи защищают растение от других насекомых и растений-паразитов. Любое насекомое, которое может убить или съесть растение, муравьи быстро отлавливают и убивают, что также идёт им на пользу[2][3][4].
Специализированные структуры растения свидетельствуют об очень интересной коэволюции и симбиозе между этими двумя видами. Недавнее исследование, проведённое Селин Леруа с соавторами (2008), выявило множество особенностей, которые растение выработало для поддержки муравьёв. Во-первых, домации расположены рядом со стеблями, которые муравьи используют для охоты. Во-вторых, они содержат внецветковые нектарники и пищевые тела, чтобы кормить муравьёв, если у них нет других источников пищи. В-третьих, внутри домаций обнаружено меньше хлоропластов, что означает, что она обладает меньшей фотосинтетической способностью. В-четвёртых, внутри домика были обнаружены стоматы, хотя и с меньшей плотностью, возможно, для улавливания углекислого газа, образующегося в результате дыхания муравьёв. Наконец, в домациях было обнаружено большее количество целлюлозы, что привело к утолщению клеточной стенки и созданию более жёсткой поверхности, способной выдержать вес муравьёв. Эти выводы показывают, что участки, предназначенные для домаций, являются наследственными и результатом коэволюции; они были созданы исключительно для того, чтобы поддерживать этот конкретный вид муравьёв[3].
Симбиоз с грибами
В домациях, где живут муравьи, растёт множество различных видов грибков. Когда королева-основательница только начинает откладывать яйца в своём гнезде в H. physophora, гифы множества различных видов грибов начинают расти, постепенно обволакивают гнездо и полностью закрывают вход в него. Когда созревают и появляются первые рабочие муравьи, им приходится пробивать себе путь сквозь грибковое покрытие, чтобы попасть на внешнюю сторону растения. Однако, как обнаружили мирмекологи, ещё более примечательно то, что из всех этих видов грибов A. decemarticulatus культивирует только один конкретный вид. Этот гриб представляет собой сажистую плесень, которая, согласно одному из исследований Марио Руис-Гонсалеса с соавторами (2010), характеризуется близкородственными гаплотипами (генетическими маркерами) в порядке Chaetothyriales из класса Эуроциомицеты. Муравьи используют эту плесень для строительства своих ловушек[5].
Мирмекологи также отметили, что в отличие от типичных мутуализмов между муравьями и грибами, эти муравьи не получают от гриба никакого питания. Они строго манипулируют физиологией гриба, чтобы построить раствор для ловушки, способной поймать гораздо более крупную добычу[5].
Изготовление ловушки
Создание ловушки — это необычное для муравьёв действие. Рабочие сооружают ловушки на стеблях растения, в основном делая приподнятую и полую платформу на одном из участков стебля. Сама конструкция выглядит как часть растения, как будто она немного увеличилась в ширину. Муравьи также проделывают в этой платформе небольшие отверстия, которые немного шире их собственных тел. Отдельные рабочие особи прячутся в этих отверстиях под поверхностью, невидимые для своей жертвы снаружи. Они будут располагать свои головы снаружи растения с открытыми мандибулами, ожидая добычу[2][4].
Изготовление ловушки происходит путём срезания растительных волосков (трихом) с узкого вертикального участка стебля за пределами домация. Затем муравьи раскладывают эти волоски, чтобы очертить структуру ловушки, и срыгивают собранную плесень, которая действует как паста и удерживает трихомы вместе. Для сбора этой плесени они собирают мицелий гриба, растущего на растении. Эта плесень будет продолжать расти между трихомами и вокруг отверстий, заполняя и укрепляя структуру[2][5].
Хищническое поведение
Обычно на стебле растения находится небольшое количество муравьёв. Изучение суточного ритма активности позволило отметить постоянное патрулирование листьев, но с глобально большим количеством рабочих в дневное время, чем в ночное, и сильным снижением в сумерках. Среднее число рабочих на лист варьировало от 0,3 между 19 и 21 часами до 2,3 в 10 часов утра. То есть, патрулирующие рабочие часто находились на листе в одиночестве, когда присутствовали на нем. Как только насекомое садится на растение, ближайший муравей высовывается из отверстия на стебле и немедленно хватается за его ногу, антенну, крыло или другой придаток добычи. Муравей потянет за собой добычу, которая попытается вырваться, потянувшись в противоположном направлении. Однако муравей редко отпускает свою хватку. Муравей обездвиживает добычу и, проявляя небывалую силу, удерживает гораздо более крупную добычу на растении, в то время как на него сбегаются другие муравьи из этого района. Эта способность удерживать добычу чрезвычайно важна, поскольку муравьи передвигаются и собираются довольно медленно[2].
После того как первый муравей обездвижил добычу, он выпускает феромоны, чтобы призвать других муравьёв в этот район. Первые несколько муравьёв хватают добычу за разные ноги и тянут в разные стороны, «растягивая» её[2]. В таком растягивании участвуют от шести до 16 муравьёв. Когда добыча лежит неподвижно и беспомощно, рабочие муравьи собираются вместе и либо помогают удерживать добычу, если это необходимо, либо начинают жалить и кусать её. При этом они используют вырабатываемый ими яд, чтобы парализовать и убить добычу. После этого группа рабочих несёт тело обратно в колонию, где его расчленяют и разрезают на мелкие кусочки, чтобы съесть всей группой[2].
Это хищническое поведение — яркий пример согласования между одиночным и совместным хищничеством. Первый муравей сначала действует в одиночку, но потом приходят другие муравьи и присоединяются к охоте, а третьи возвращаются в домации, чтобы мобилизовать ещё больше муравьёв для помощи в удержании и последующем расчленении добычи. Это также демонстрирует, насколько эффективнее может быть группа особей, чем одиночка, работающий сам по себе. Одиночный муравей никогда не смог бы убить такую крупную добычу; они способны сделать это только благодаря коммуникации и сотрудничеству[2].
Охота на кузнечиков и термитов
Такое хищническое поведение очень эффективно в отношении относительно крупной добычи, которую потребляют муравьи. Дежан и другие исследователи (2001) проверили пределы его эффективности, подвергнув муравьёв воздействию термитов Nasutitermes (длина около 6 мм) и кузнечиков (около 1 см), которые примерно в 40 и 142,2 раза больше рабочих муравьёв, соответственно. Таким образом, один рабочий Allomerus decemarticulatus, сдерживающий кузнечика, примерно эквивалентен человеку весом 80 кг, удерживающему объект весом около 11 тонн, который активно пытается вырваться. Когда термиты были предложены для эксперимента, каждый из них был схвачен муравьями; ни один не смог убежать. Однако в случае с кузнечиками все они были схвачены по крайней мере за одну ногу, но из 20, предложенных муравьям, пятеро были схвачены полностью, 12 убежали, но потеряли ногу, за которую ухватился рабочий, а семеро ускакали[2].
Любая добыча, обладающая способностью прыгать или летать, всегда будет спасаться более эффективно, чем нелетающие насекомые, такие как термиты. Однако во многих случаях те, кому удаётся спастись, все равно теряют ногу или другой придаток, как 60 % кузнечиков в эксперименте. Поскольку добыча намного крупнее муравьёв, даже получение одного придатка в качестве источника пищи — уже победа. Например, задняя нога кузнечика все равно примерно в 12,4 раза больше одного муравья и сама по себе является большим ресурсом[2].
Охота на термитов отличается почти 100%-й результативностью. Размер рабочих A. decemarticulatus и их очень медленное передвижение не позволили исследователям отметить, была ли добыча обнаружена при контакте или с небольшого расстояния. Рабочие, которые никогда не выставляли антенны на встреченных термитов, сразу же хватали их за ногу, затем тянули назад, в то время как добыча тянулась в противоположном направлении. Такая ситуация сохранялась до прихода новых патрулирующих муравьёв (от 4 до 40 минут; 19,75±3,4 минуты; N =12). Муравьи, пришедшие «вторыми», всегда захватывали другую ногу, затем тянули назад, тем самым начиная растягивать добычу. Прибывшие «третьими», «четвёртыми» и «пятыми» делали то же самое в 85,7 %, 53,6 % и 10,7 % случаев, соответственно (N = 28). Оставшиеся рабочие возвращались в домации, чтобы провести мобилизацию соплеменников на большом расстоянии. Затем поочерёдно прибывали новые патрулирующие муравьи, а также завербованные члены их гнезда. Первые прибывшие также захватывали ногу добычи и тянули её назад, в результате чего добычу удерживали от шести до 16 муравьёв. Среди муравьёв, прибывших позже, одни жалили и кусали добычу, а другие патрулировали территорию. Благодаря совместной транспортировке добыча была извлечена целиком, а общая продолжительность захвата варьировала от 10 минут до часа. Все испытанные термиты были пойманы[2].
Сравнение с другими муравьями
Хотя стратегия патрулирования A. decemarticulatus в основном одиночная, хищническое поведение полностью отличается от одиночной охоты дендробионтных муравьёв, которые осваивают добычу в одиночку, даже достаточно крупную, чтобы её не мог перенести один рабочий. Охота у A. decemarticulatus зависит от того, что одиночные рабочие способны обездвиживать добычу в течение длительного времени, позволяя другим муравьям прибывать последовательно, причём первые участвуют в растягивании добычи, а прибывающие позже дополнительно вербуют соплеменников на большом расстоянии. В результате добыча оказывается растянутой, что напоминает поведение доминирующих дендробионтных муравьёв (то есть видов с большими колониями, чьи территории распределены мозаично в лесных пологах из-за внутривидовой и межвидовой агрессивности). Как и муравьи-ткачи Oecophylla longinoda, которые никогда не используют яд, A. decemarticulatus захватывают добычу целиком, и как Crematogaster, которые разделывают добычу на месте, A. decemarticulatus используют яд. Тем не менее, доминирующие муравьи добывают пищу группами, что позволяет им быстро мобилизовать рабочих муравьёв на короткие расстояния, когда она начинает захватывать добычу. Они используют дальнюю вербовку только для очень крупной добычи, для освоения которой и последующего извлечения требуется большое число рабочих. Исследователи не исключают существование ближней мобилизации у A. decemarticulatus, когда два или более рабочих патрулируют на одном листе[2].
Взаимодействие с другими животными
Если многочисленные взаимоотношения A. decemarticulatus не были достаточно сложными с другими насекомыми, они также часто взаимодействуют с клопом-хищнецом Zelus annulosus, который часто обитает на растениях H. physophora. Однако эти клопы имеют адаптированные физиологические и поведенческие характеристики, которые позволяют им избегать хищничества A. decemarticulatus, сохраняя при этом мутуалистические отношения с растением[6].
Подобно муравьям, Z. annulosus обычно живёт на молодых особях H. physophora, где самки откладывают яйца на стебель. Когда они начинают развиваться, молодые клопы живут среди трихом стебля и охотятся на листьях растения. Отношения между клопом-хищнецом и растением действуют независимо от отношений между муравьями и растением. Однако разница между этими двумя отношениями заключается в том, что клопы-хищнецы не берут у растения источник пищи, как это делают муравьи[6].
Z. annulosus в основном приспособился жить и охотиться вокруг A. decemarticulatus. Клопы-хищнецы используют именно это растение для выращивания своих нимф, поскольку трихомы растения отпугивают более крупных муравьёв, которые могут убить молодых развивающихся клопов. Кроме того, клопы выделяют липкое вещество, которое позволяет им ходить по трихомам, избегая ловушек A. decemarticulatus. Таким образом, в отношениях между Z. annulosus и H. physophora клоп получает укрытие от потенциальных крупных хищников-муравьёв, а растение — вторую линию защиты от травоядных. Клоп-хищнец также мирно уживается на растении с муравьями. Они охотятся в сходных местах на растении, но клопы-хищнецы, как предполагается, активно избегают муравьёв, поскольку они гораздо быстрее муравьёв[6].

Таксономия
Вид был впервые описан в 1878 году австрийским мирмекологом Густавом Майром (Gustav Mayr) по типовому материалу рабочего муравья из Бразилии[7]. Муравьиная матка (королева) была впервые кратко описана в 1975 году бразильско-американским энтомологом и монахом-францисканцем Вальтером Кемпфом (Walter W. Kempf, 1920—1976)[8], а более подробно в 2007 году колумбийским мирмекологом Фернандо Фернандесом (Богота, Национальный университет Колумбии, Колумбия) в ходе его ревизии рода Allomerus. Самцы неизвестны[1].
Распространение
Эндемик неотропической Южной Америки. Встречаются в Бразилии и Французской Гвиане[1]. Этот вид обитает на Hirtella physophora (из семейства двудольных растений Хризобалановые), ареал которого ограничен Бразилией и Французской Гвианой[4]. Они живут и охотятся на H. physophora, живя в мешочках на листьях и патрулируя стебель растения в поисках добычи[3].
Примечания
- 1 2 3 4 5 6 Fernandez F. The Myrmicine ant genus Allomerus Mayr (Hymenoptera: Formicidae) (англ.) // Caldasia : Журнал. — Instituto de Ciencias Naturales, Universidad Nacional de Colombia, 2007. — Vol. 29. — P. 159–175. — .
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Dejean A., Solano P. J., Belin-Depoux M., Cerdan P., Corbara B. Predatory behavior of patrolling Allomerus decemarticulatus workers (Formicidae: Myrmicinae) on their host plant (англ.) // Sociobiology : Журнал. — 2001. — Vol. 37. — P. 571–578.
- 1 2 3 4 Leroy C., Jauneau A., Quilichini A., Dejean A., Orivel J. Comparison between the anatomical and morphological structure of leaf blades and foliar domatia in the ant-plant Hirtella physophora (Chrysobalanaceae) (англ.) // Annals of Botany : Журнал. — 2008. — Vol. 101. — P. 501–507. — doi:10.1093/aob/mcm323. — PMID 18222908. — PMC 2710197..
- 1 2 3 Grangier J., Dejean A., Malé P., Orivel J. Indirect defense in a highly specific ant-plant mutualism (англ.) // Naturwissenschaften : Журнал. — 2008. — Vol. 95. — P. 909–916. — doi:10.1007/s00114-008-0398-4. — . — PMID 18496661.
- 1 2 3 Ruiz-González M., Malé P. G., Leroy C., Dejean A., Gryta H., Jargeat P., Quilichini A., Orivel J. Specific, non-nutritional association between an ascomycete fungus and Allomerus plant-ants (англ.) // Biology Letters : Журнал. — 2010. — Vol. 7. — P. 475–479. — doi:10.1098/rsbl.2010.0920. — PMID 21084334. — PMC 3097849.
- 1 2 3 Revel M., Dejean A., Céréghino R., Roux O. An assassin among predators: the relationship between plant-ants, their host Myrmecophytes and the Reduviidae Zelus annulosus (англ.) // PLoS ONE : Журнал. — 2010. — Vol. 5. — doi:10.1371/journal.pone.0013110. — . — PMID 20957040. — PMC 2948506.
- ↑ Mayr G. Formiciden gesammelt in Brasilien von Professor Trail (нем.) // Verh. K-K. Zool.-Bot. Ges. Wien : Журнал. — 1878 (1877). — Vol. 27. — P. 867–878. (page 874, описание рабочего муравья)
- ↑ Kempf W. W. Miscellaneous studies on neotropical ants. VI. (Hymenoptera, Formicidae) (англ.) // Stud. Entomol. : Журнал. — 1975. — Vol. 18. — P. 341–380. (page 347, описание самки-королевы)
Литература
- Dejean A., Solano P., Ayroles J., Corbara B., Orivel J. Arboreal ants build traps to capture prey (англ.) // Nature : Журнал. — Springer Nature Limited, 2005. — Vol. 434, no. 973. — P. 673–675. — doi:10.1038/434973a.
- Malé P.-J. G., Loiseau A., Estoup A., Quilichini A., Orivel J. Characterization of polymorphic microsatellite loci in the neotropical plant-ant Allomerus decemarticulatus (Formicidae: Myrmicinae) and multiplexing with other microsatellites from the ant subfamily Myrmicinae (англ.) // European Journal of Entomology : Журнал. — 2010. — Vol. 107, no. 4. — P. 673–675. — doi:10.14411/eje.2010.074.
- Solano P.-J., Durou S., Corbara B., Quilichini A., Cerdan P., Belin-Dupoux M., Delabie J.H.C., Dejean A. Myrmecophytes of the understory of French Guianian rainforests: their distribution and their associated ants (англ.) // Sociobiology : Журнал. — 2003. — Vol. 41, no. 2. — P. 605–614.
Ссылки
- Мураши ловят монстров в живые капканы и разводят грибы (англ.). Мембрана. Дата обращения: 15 января 2025. Архивировано 12 мая 2011 года.
- Species: Allomerus decemarticulatus (англ.). antweb.org. Дата обращения: 15 января 2025.
- Allomerus decemarticulatus Mayr, 1878 (англ.). Biolib.cz. Дата обращения: 15 января 2025.